歡迎來到高新區數耕網絡科技工作室-專業無紡布袋定制、批發生產廠家! 381708881@qq.com 18482170382 |


對于治療嚴重的心臟瓣膜疾病,人工瓣膜在大多數成年人中效果相當好;然而,對于兒科患者,還有一個額外的要求,即更換瓣膜要隨著孩子一起成長,因此極大地限制了目前的治療選擇。組織工程心臟瓣膜 (TEHV),例如源自自體骨髓干細胞 (BMSC) 的瓣膜,有可能重現天然瓣膜結構并適應軀體生長。然而,促進與天然組織的定向整合而不是隨機、不受控制的生長的基本前提是了解 BMSC 對瓣膜相關機械環境的機械生物學反應。在這里,我們報告了人類 BMSC 接種聚合物結構對瓣膜相關應力狀態的反應:(i) 單獨穩定流動,(ii) 單獨周期性彎曲,以及 (iii) 周期性彎曲和穩定流動 (彎曲流動) 的組合。將 BMSC 接種到 PGA:PLLA 聚合物支架上并在靜態培養中培養 8 天。隨后,再施加上述機械條件(組別包括穩定流(850ml/min)、周期性彎曲(1 Hz)和彎曲流(850ml/min 和 1 Hz)。我們發現彎曲流組的樣本呈現瓣膜樣細胞分布,這些細胞表達內皮細胞(偏向表面)和肌成纖維細胞(偏向中間區域)表型。我們認為這可能是由于存在明顯的流體誘導剪切應力大小和振蕩剪切應力,這些剪切應力同時施加在樣本上。這些結果表明,彎曲流機械環境支持 BMSCs 體外定向分化為心臟瓣膜表型,這可通過細胞分布和特定基因標記的表達來證明。在柔性流動條件下,BMSC 衍生的工程組織生長的先驗指導可能有助于隨后促進體內受控的、工程化的天然組織整合過程,而這對于成功實現長期瓣膜重塑必不可少。
一、簡介
心臟瓣膜在控制單向血流方面起著重要作用。然而,出生缺陷或感染(例如風濕熱)可能導致一個或多個心臟瓣膜功能障礙,這可能導致兒童出現嚴重的瓣膜異常。植入物設計和手術技術的進步大大提高了成人患者人工心臟瓣膜的成功率。然而,由于這些植入物無法促進軀體生長和瓣膜組織重塑,因此其在兒科患者中的療效受到嚴重限制;因此,多次大手術和再次手術很常見,這給成長中的孩子帶來了巨大的健康負擔。
與天然瓣膜類似,組織工程心臟瓣膜 (TEHV) 具有適應和隨宿主進化的能力,在概念上被認為是治療心臟瓣膜疾病的永久解決方案。在每個心動周期中,天然瓣膜都會因血流而不斷受到機械應力;例如,主動脈瓣葉在收縮中期會經歷由液體引起的峰值剪應力,約為 5-6 達因/厘米2。機械刺激應用于發育中的心血管組織時,會改變基因表達,促進組織重塑事件,進而增強特定的機械和表型特征。天然心臟瓣膜會受到高度復雜的周期性、拉伸和液體引起的應力。在工程心臟瓣膜組織中,機械刺激,尤其是結合液體引起的剪應力的刺激,可增強祖細胞分化途徑并構建用于瓣膜應用的組織特性。例如,研究人員使用脈動流生物反應器裝置進行實驗,該裝置增強了 TEHV 三葉結構,與靜態培養的對應物相比,膠原蛋白細胞外基質 (ECM) 含量增加了約 300%。在其他地方,已經建造了生物反應器來耦合任何組合的流動、循環拉伸和循環彎曲 (FSF 生物反應器),這也證實了耦合的機械刺激顯著促進 ECM 的產生;特別是,與心臟瓣膜相關的穩定流與循環彎曲的組合,研究人員表明,血流誘導的振蕩剪切應力 (OSS) 直接調節斑馬魚模型中來自 kruppel 樣因子基因家族 KLF2A 的轉錄因子的正常表達。KLF2A 基因與瓣膜形成密切相關,其缺失會導致心臟瓣膜缺陷。
由于缺乏有關優化體外培養過程所需的機械生物學事件的信息,功能性 TEHV 的生成仍然難以捉摸。盡管如此,在機械指導下,組織工程可行性研究迄今已證明,天然瓣膜細胞可以重現具有足夠機械強度和形態的瓣膜結構。此外,非瓣膜細胞,如骨髓干細胞 (BMSC)、隱靜脈內皮細胞 (EC)、升主動脈肌成纖維細胞和臍帶衍生細胞,在機械調節狀態下表現出瓣膜 ECM、DNA 含量和體外內皮化的增加。
BMSC 尤其在心臟瓣膜組織工程方面顯示出巨大的前景,因為它們是多能干細胞,免疫原性風險極小,沒有道德/法律問題,并且可以輕松獲取和培養擴增;通常,BMSC 可以在幾天內分離、純化并大量擴增。BMSC 在體外保持廣泛的分化、增殖和克隆形成能力。人類 BMSC 對機械調節有反應,并且已被證明可以在體外產生心臟瓣膜 ECM 成分。研究人員描述了闡明組合循環彎曲和穩定流動狀態 (flex-flow) 的影響的開創性工作,這有助于顯著促進源自 BMSC 的從頭工程心臟瓣膜組織中的工程膠原蛋白。然而,在組織規模上評估 TEHV 研究的一個基本前提是需要了解瓣膜相關機械刺激可以調節細胞命運的過程,特別是在干細胞來源的背景下的細胞分化。因此,在這項研究中,我們的主要目標是確定 BMSC 對 flex-flow 條件在促進瓣膜形成方面的獨特反應。
二、材料與方法
2.1 BMSCs 培養與擴增
從人骨髓中分離的 BMSCs,用基質、干細胞和造血標志物進行表征和測試,購買于冷凍小瓶中。從干冰包裝中回收細胞,立即解凍并用預熱的培養基稀釋,以減少到達時冷凍保存試劑的毒性作用。將約 5 x 10^5 個細胞轉移到 T75 通風細胞培養瓶中,并置于 37℃、濕度為 95%、CO2 為 5% 的培養箱中。培養瓶在 1 周時完全匯合。每次傳代平均獲得 2 x 10^6 個細胞。使用新鮮配制的干細胞培養基(含 10% 高級干細胞未分化生長補充劑、1% 青霉素和鏈霉素培養基)進行細胞培養與擴增。在整個實驗過程中,我們一直保持無菌細胞和組織培養環境。第 6 至 8 代的 BMSC 用于后續的組織工程實驗。
2.2 支架制備和細胞接種
支架材料為等比例的聚乙醇酸 (PGA) 和聚 L-乳酸 (PLLA) 無紡布聚合物氈 。切割長 17 毫米、寬 6 毫米、厚 1 毫米的樣本 (n = 12),并將兩個金屬彈簧小心地連接到支架的兩端。在我們定制的 U 形生物反應器裝置中進行動態組織培養需要此組件 (圖 1);更多圖紙和細節可參見研究人員的文章。在細胞接種之前,用環氧乙烷 (EtO; AN 306, Anprolene, Andersen Products Inc, HawRiver, NC) 對支架進行氣體滅菌 12 小時,并用 70% 乙醇處理。按照制造商的說明指南中所述的建議進行曝氣程序。使用以下方案完成接種程序:用杜氏磷酸鹽緩沖鹽水 (DPBS, Fisher Scientific) 緩沖液沖洗約 90% 的匯合燒瓶。接下來,加入 0.25% 胰蛋白酶和乙二胺四乙酸 (EDTA) 溶液,在 37℃ 下孵育 3 分鐘。用等體積的血清中和胰蛋白酶溶液。將細胞懸浮液收集在 15 毫升錐形管中,以 1700 rpm 的速度離心 5 分鐘。通過去除上清液回收細胞沉淀,并用新鮮制備的組織培養基懸浮以供進一步使用。單個支架上接種了 2 x 10^6 BMSC,并懸浮在 50 毫升通風錐形管中的 20 毫升組織培養基中。組織培養基由杜爾貝科改良的 Eagle 培養基 (DMEM,Fisher Scientific) 組成,其中補充了 10% 胎牛血清、1% 青霉素和鏈霉素、2ng/ml 堿性成纖維細胞生長因子和 82 μg/ml 抗壞血酸 2 磷酸鹽 (AA2P,Sigma-Aldrich)。隨后,將這些管放入細胞和組織培養箱內的旋轉式烤盤中,轉速為 8 RPM。每兩天更換一次培養基,并將 BMSC 接種的支架在旋轉式烤盤培養下培養,總共 8 天。

東莞市富臨塑膠原料有限公司 是 PLGA Biofelt植入級無紡布的專家, 富臨塑膠為客戶提供“服務”和“PLGA Biofelt無紡布”。
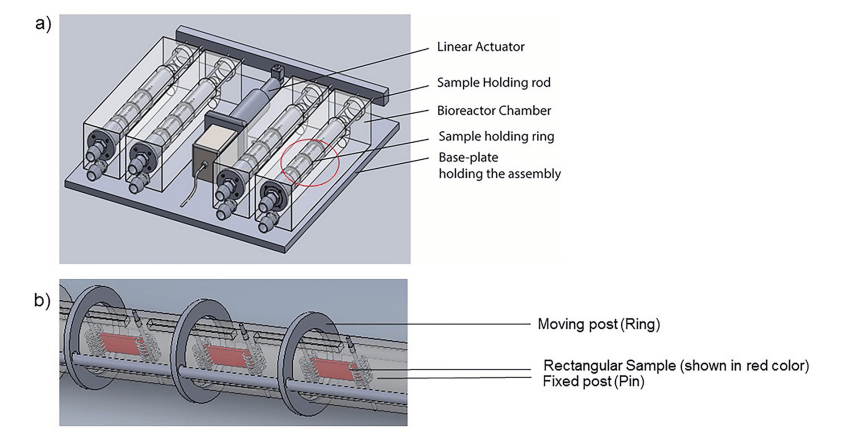
圖 1. a) 定制 U 形生物反應器的示意圖,該生物反應器連接到線性致動器,該致動器引導穿過樣品的桿,使它們彎曲和伸直。b) 插圖:顯示調節室內的三個樣品,一端(環)可以移動,另一端(銷)固定。在當前研究中,致動器設置為 1 Hz 頻率以允許周期性彎曲,同時泵以 850 ml/min 的穩定流速運行。
2.3 組織工程實驗
在前 8 天的旋轉培養后,將樣本隨機分配到四個不同的處理組之一,并進行額外的 14 天組織培養。這些組為(n = 12 個樣本/組):1) 無流動(靜態控制),2) 單獨穩定流動(流動),3) 單獨樣本循環彎曲(彎曲)和 4) 穩定流動和樣本循環彎曲的組合(彎曲流動)。對于機械調節組,使用之前詳細描述過的定制 U 形生物反應器。該裝置連接到蠕動泵和環境密封的線性執行器以用于實驗目的。實驗裝置由四個相同的調節室組成,每個室包含 3 個樣本。標本的一端用銷釘固定,另一端連接到圓形移動柱上。對于 Flex 和 Flex-Flow 實驗,移動柱沿軸向線性移動,使用線性致動器(頻率為 1 Hz)啟動樣品循環彎曲。對于 Flow 和 Flex-Flow 案例,使用蠕動泵來維持 850 ml/min 的連續流速(S1 文件)。
2.4 膠原蛋白含量
所有四組的樣本在 22 天后取出(每組 n = 3)并進行膠原蛋白生化測定。消化的膠原蛋白樣品的定量方法與之前描述的方法類似。用 0.5 M 乙酸溶液(Sigma)和胃蛋白酶(1 mg/ml 胃蛋白酶(P7000),Sigma)消化樣品。在搖床上在 4℃ 下進行 16 小時的消化。然后根據 Sircol 可溶性膠原蛋白測定試劑盒(Biocolor Ltd.)提供的體外組織程序對膠原蛋白提取物進行測定。將多模式微孔板讀數儀設置為 555 nm 的吸光度以獲得膠原蛋白濃度。
2.5 組織學
為了檢測關鍵瓣膜 ECM 成分(膠原蛋白、糖胺聚糖 (GAG) 和彈性蛋白)的存在,對培養的工程組織和新鮮收獲的豬天然主動脈瓣葉(作為陽性對照)進行了組織學處理。在室溫下,用 DPBS 清洗樣品,并用 10% 福爾馬林固定過夜,體積分數為 20:1。固定后,用剪刀剪下 2 毫米 x 2 毫米的組織切片,并嵌入組織冷凍培養基中。為了保留結構,將組織在室溫下用液體冷凍培養基在培養皿中孵育 2 小時。接下來,還將液體冷凍培養基單獨添加到塑料模具中,并在液氮溫度下冷凍。約 1 分鐘后,獲得白色固體冷凍介質床。此時,將準備好的組織切片小心地放在模具內的冷凍固體床上,并添加更多冷凍介質以浸泡樣品。隨后將模具在液氮中速凍。最后,使用低溫恒溫切片切割與組織表面平行的連續切片(厚度為 12 μm)。將切片安裝到玻璃載玻片上。使用制造商提供的染料將組織學染色劑應用于組織切片,以便隨后進行顯微鏡可視化。
2.6 免疫熒光染色
22 天后對 CD31 和 α-SMA 細胞表面蛋白進行免疫熒光染色(n = 3 個樣本/組),以分別檢測內皮和肌成纖維細胞表型。本研究重點關注兩種瓣膜相關蛋白,例如分化簇 31 (CD 31) 和 α-平滑肌肌動蛋白 (α-SMA)。已知 CD31 表達于 EC 表面和 SMC 中的 α-SMA,以及瓣膜間質細胞中的肌成纖維細胞樣細胞。每個樣本沿組織切片,用抗 αSMA 和抗 CD31 對三個不同層(即頂部表面層、深層中核層和底部表面層)進行免疫染色,并評估 SMC 和 EC 表達情況。解剖從心臟分離的豬主動脈瓣膜并將其指定為陽性對照。按照相同的方案對瓣膜小葉進行免疫染色以檢測 EC 和 SMC 表達。
樣品固定、包埋和切片程序與前面描述的組織學方案相同(“材料和方法”中的“組織學”子部分)。接下來,對于免疫熒光檢測,遵循以下染色程序:將安裝在玻璃載玻片上的組織切片用 0.1% Triton X-100 處理 3 至 5 分鐘以增強細胞質的通透性(CD31 染色不包括此步驟)。用 DPBS 進行額外的洗滌步驟三次,每次 5-10 分鐘。通過在 DPBS 中加入 1% 山羊血清 30 分鐘來促進非特異性表位的阻斷。所用的一抗為小鼠單克隆抗 CD31 和小鼠單克隆抗 α SMA。在 4℃ 下孵育過夜。用洗滌緩沖液 (DPBS+ 0.01% Triton-X 100) 洗滌樣品以降低背景。加入二抗 (山羊多克隆抗小鼠 IgG (H+L) (DyLight 488) (Fisher Scientific) 含 1% 山羊血清),并在 4℃ 下孵育過夜,以進行 CD31 和 α-SMA 免疫熒光染色。在熒光顯微鏡下觀察帶有染色切片的玻璃載玻片。對于 CD31,距離表面頂部 92±10.58 μm 的深度被視為頂層,距離底部表面 76±4 μm 的深度被視為底層,距離頂部表面 264±18.33 μm–652±17.43 μm 的深度被視為深層中芯層。同樣,對于 α-SMA,距離頂部表面 ~140±17.43 μm 的深度被視為頂層,距離底部表面 ~108±12 μm 的深度被視為底層,距離頂部表面 336±13.85 μm–464±21.16 μm 的深度被視為深層中芯層。
2.6.1 圖像分析
使用以下協議 (ImageJ, NIH; Bethesda, MD) 量化 CD31 和 α-SMA 染色圖像 (樣本大小, n = 3) 的信號強度值。定義一個面積為 10,000 像素的矩形感興趣區域 (ROI)。選擇并測量三個具有最大強度的不同 ROI (重復, r = 3)。ROI 的平均強度以任意單位 (AU) 記錄。注意避免任何假陽性信號。
2.7 定量實時聚合酶鏈式反應 (QRT-PCR)
組織培養 22 天后,對四組新生組織進行基因表達評估,并對有限數量的瓣膜相關標志物進行量化,以驗證免疫染色結果。如前所述,進行定量實時聚合酶鏈式反應 (QRT-PCR)。用 DPBS 清洗工程組織,并根據制造商的方案 (SV Total RNA Isolation System,Promega) 分離總 RNA,從小組織樣本 (≤30mg) 制備裂解物。簡而言之,將 1ml 裂解緩沖液加入 30 mg 組織中,然后快速冷凍并用均質機高速均質,直至混合物中沒有可見的組織碎片。將 175 μl 組織裂解物轉移到 1.5 ml 微量離心管中。將沉淀物轉移至 1.5ml 無 RNase 微量離心管中進行 RNA 分離。用 SV 總 RNA 分離系統 (Promega) 純化總 RNA。利用分光光度計 (Varian Carry 300) 驗證分離的 mRNA 濃度和質量。所有樣品測量均準備了 60 倍稀釋液。RNA 純化數據在補充數據表 (S3 文件) 中提供。1 μg 總 RNA 用于逆轉錄反應,并使用 GoScript 逆轉錄系統 (Promega) 提供的寡核苷酸 (dT) 15 引物合成 cDNA。使用 GoTaq_qPCR Master Mix (Promega) 進行 QRT-PCR。用 Step-One 實時 PCR 系統 (Applied Biosystems) 檢測信號強度。PCR 混合物包含正向和反向引物以及 SYBR 綠 I 染料試劑,以及從逆轉錄中獲得的 cDNA。引物和基因均選自參考來源,所有引物均購買(Sigma Aldrich)。引物序列(表 1)GAPDH、YARS、KLF2A 序列使用 BLAST 程序獲得,國家生物技術信息中心 (NCBI),FZD2 和 MLC1F 來自研究人員。簡而言之,實驗所用的條件如下:PCR 管在 95℃ 下保持 2 分鐘,然后開始循環以激活 Taq 聚合酶。循環參數為 95℃ 持續 5 秒;60℃ 持續 45 秒;95℃ 持續 15 秒。使用試劑指南中描述的 ΔΔCt 方法,對閾值循環 (ΔCt) 值的變化取平均值,并用內源基因 GAPDH 進行標準化。基因倍數變化的表達計算為 2- ΔΔCt,并比較四組(Static、Flow、Flex 和 Flex-Flow、PHV)的基因表達率以進行進一步分析。在每個完整的 QRT-PCR 循環后進行熔解曲線分析,以檢查任何污染、引物質量或重復擴增。GAPDH、YARS、FZD2、MLC1F、KLF2A 的熔解溫度 (Tm) 分別記錄為 (82.6±0.001)℃、(80.05±0.007)℃、(81.97±0.08)℃、(79.91±0.09)℃ 和 (79.92±0.001)℃。每組的熔解曲線在補充數據(S4 文件)中提供。
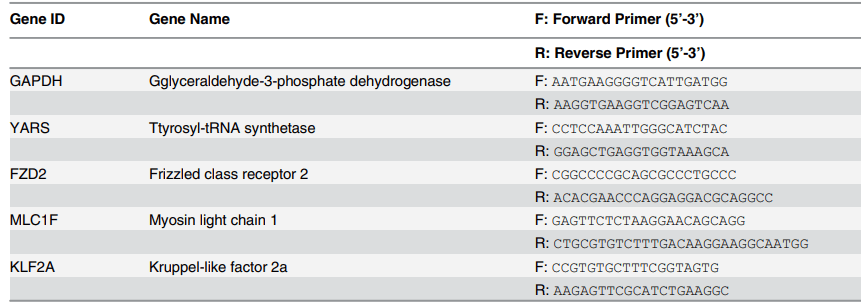
表 1. 本研究中用于 RT-PCR 分析的引物序列。
2.8 計算流體動力學 (CFD)
為了量化施加在生物反應器樣品上的流體誘導應力,我們進行了 CFD 模擬。對于單獨的彎曲流動和循環彎曲的情況,我們結合了移動邊界分析;我們之前的工作詳細描述了 CFD 方法的細節。模擬了單獨流動、單獨循環彎曲、流動和彎曲組合以及無流動四種情況。對于單獨的彎曲流動和穩定流動的情況,我們使用了 0.1067 m/s 的入口速度邊界條件,這代表了實驗規定的 850 ml/min 的流速。生物反應器壁沒有規定滑移條件,而對于單獨的彎曲流動和循環彎曲,樣品壁的速度設置為等于網格速度。生物反應器的出口設置為零相對壓力邊界條件。所有模擬均采用牛頓粘性模型,采用層流條件,流體材料特性如下:密度 = 1.01g/cm3,動態粘度 = 1.27 cp。在滿足每個動量、連續性和網格位移方程的 1x10^(-9) 收斂標準后,對結果進行分析。所有模擬均在 Hewlett Packard 工作站上進行,工作站配備英特爾 (R) Xeon(R) CPU、x5550@ 2.67GHz(2 個處理器),安裝內存為 16.0 GB,操作系統為 64 位 Windows 7。
為了表示剪切應力大小和流動中時間振蕩的耦合效應,我們使用了振蕩剪切指數 (OSI) 縮放的剪切應力大小,我們之前將其定義為:


其中 TSSM 是時間平均剪切應力大小。OSI 本身定義為:
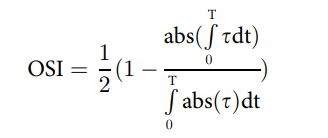
其中,“τ”是流體引起的剪切應力,“T”是周期,“t”是時間。OSI 范圍從 0 到 0.5;OSI 為 0 表示單向流動,而 OSI 為 0.5 表示流動中存在高度的時間振蕩(S5 文件)。
2.9 統計分析
進行單因素方差分析,以檢驗四組膠原蛋白生成、圖像分析和基因表達結果之間的顯著差異(n = 3 個樣本/組/結果)。隨后進行 Tukey 事后檢驗,以確定各組之間的顯著差異。所有統計分析均使用社會科學統計軟件包 (SPSS) 軟件進行。觀察到各組之間的顯著差異發生在顯著性水平 p < 0.05。
三、結果
3.1 膠原蛋白含量
生理相關的柔性流動機械調節進一步增強了工程 ECM 中的膠原蛋白:膠原蛋白是心臟瓣膜組織中的關鍵結構蛋白,在周期性彎曲、周期性拉伸和脈動流機械狀態下不斷重塑。針對心臟瓣膜的組織工程,我們研究了生理相關尺度下的彎曲和/或穩定流動模式的機械刺激如何促進 BMSCs 衍生的新生組織中的膠原蛋白含量。
靜態組的平均膠原蛋白產量為 40.91±4.23(無流動、無彎曲)、93.29±12.6(僅流動)、69.75±2.68(僅周期性彎曲)、207.38±36.61(柔性流動)μg/g 濕重(圖 2)。與其他三組相比,flex-flow 組的膠原蛋白含量明顯較高 (p<0.05)。靜態組、僅流動組和僅 flex 組之間沒有顯著差異。這一觀察結果與先前的研究結果一致。為了進行比較,評估了豬主動脈瓣葉的膠原蛋白含量 (n = 3 個瓣葉),結果為 2141.17±491.56 μg/g 濕重 (S2 文件)。
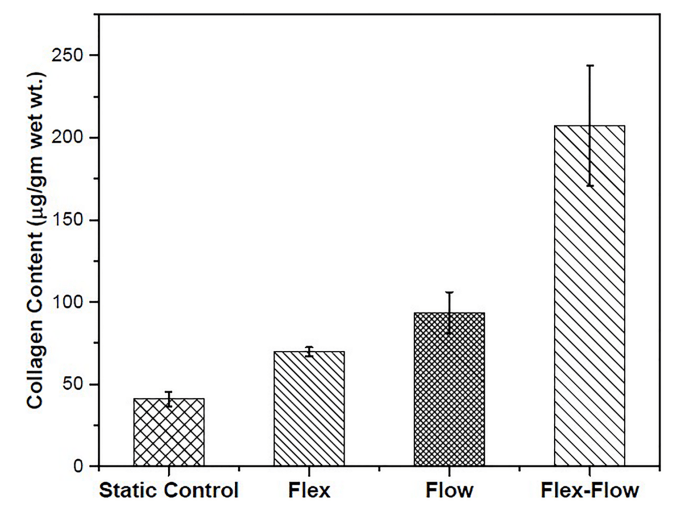
圖 2. 所研究各組樣本中的膠原蛋白含量。與所有其他組相比,Flex-Flow 組產生的膠原蛋白明顯較高 (p < 0.05)。
3.2 組織學
BMSC 植入支架的 Flex-Flow 調節可增加 ECM 中膠原蛋白和 GAG 的存在:暴露于瓣膜相關機械刺激的 BMSC 衍生工程組織經過組織學處理,以可視化瓣膜基質的三個關鍵 ECM 成分:膠原蛋白、GAG 和彈性蛋白。
經過 8 天的靜態培養后,在生物反應器中接受 Flex-Flow 機械調節 14 天的樣本顯示出工程組織 ECM 中存在膠原蛋白和 GAG 的證據(圖 3)。然而,彈性蛋白的存在尚不確定,很可能不存在,這通常是體外生長的瓣膜組織的情況。
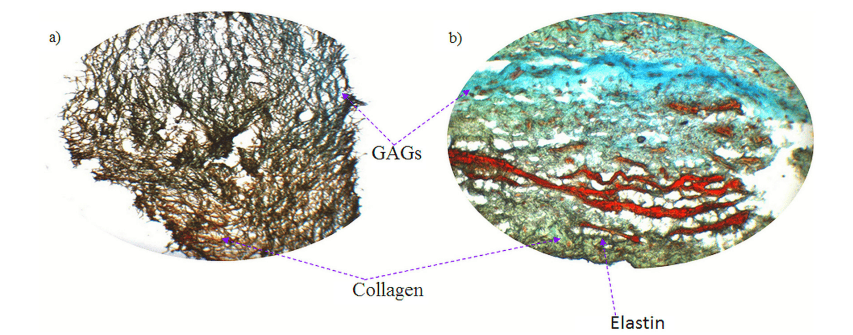
圖 3. Russell 的 Movat 五色組織學染色:a) 培養 22 天后 BMSC 接種支架的全厚度切片(8 天靜態 + 14 天 Flex-Flow 調節,b) 天然豬主動脈瓣葉的全厚度切片(+ve 對照)。顏色代碼藍色:糖胺聚糖 (GAG),黃綠色:膠原蛋白,黑色:彈性蛋白,紅色:肌肉,天然瓣膜 ECM 的三個主要成分中,膠原蛋白和 GAG 在組織工程結構中清晰可見;然而,彈性蛋白的存在尚不確定,可能不存在。
3.3 免疫熒光染色
Flex-Flow 條件促進 BMSC 分化的內皮細胞和平滑肌細胞在工程組織中的瓣膜樣分布:為了評估分化的 BMSC 在動態培養后如何在新生 ECM 中分布,對從表面到結構內更深層的工程組織連續切片進行了免疫染色。
僅在 flex-flow 組中,與瓣膜小葉表面相比,α-SMA 在中間核心層中顯著(p < 0.05)(圖 4a 和 4b)。此外,與標本的中間核心區域相比,僅在 flex-flow 組中,表面層(頂部和底部)中內皮標志蛋白 CD31 豐富(p < 0.05)(圖 5)。所有其他組均未產生 CD31 和 α-SMA 的表達(靜態、僅彎曲組)或隨機分布(僅流動)(S2 文件)。
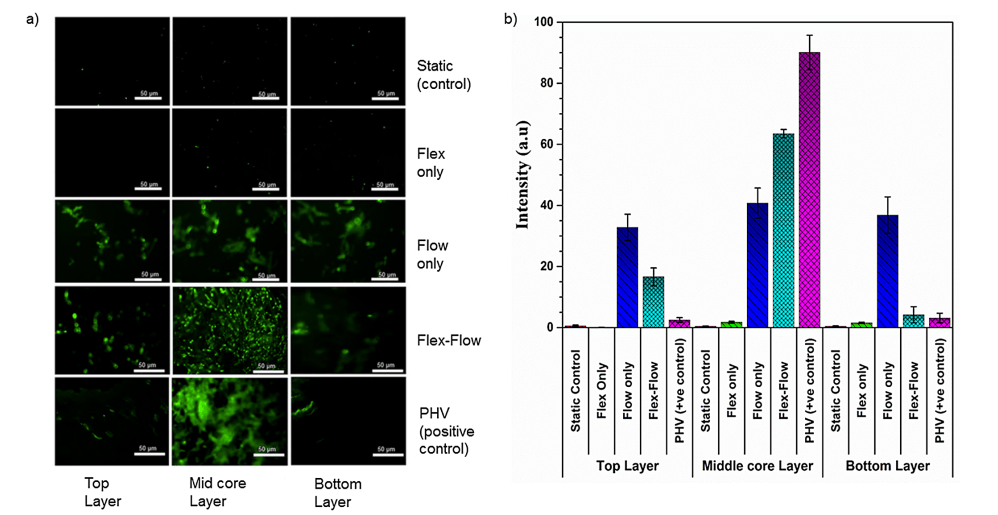
圖 4. a) 瓣膜兩個表面層(每側厚度約 90μm)、中間核心(間質組織)區域(厚度約 400 μm)上的 α-SMA 蛋白的免疫熒光染色;第一行:靜態對照;第二行:Flex;第三行:Flow;第四行:Flex-Flow 條件作用;第五行:豬心臟瓣膜作為陽性對照。在實驗組中,僅在 Flex-Flow 組中,發現 α-SMA 表達細胞在工程組織間質區域(中間層)內占主導地位;b) 量化四個實驗組中 α-SMA 信號強度的陽性染色(綠色;來自部分 a 中的圖像);與對照組相比,暴露于 flex-flow 的樣品表達的陽性 α-SMA 水平明顯更高(p < 0.05)。將 PHV 視為陽性對照。
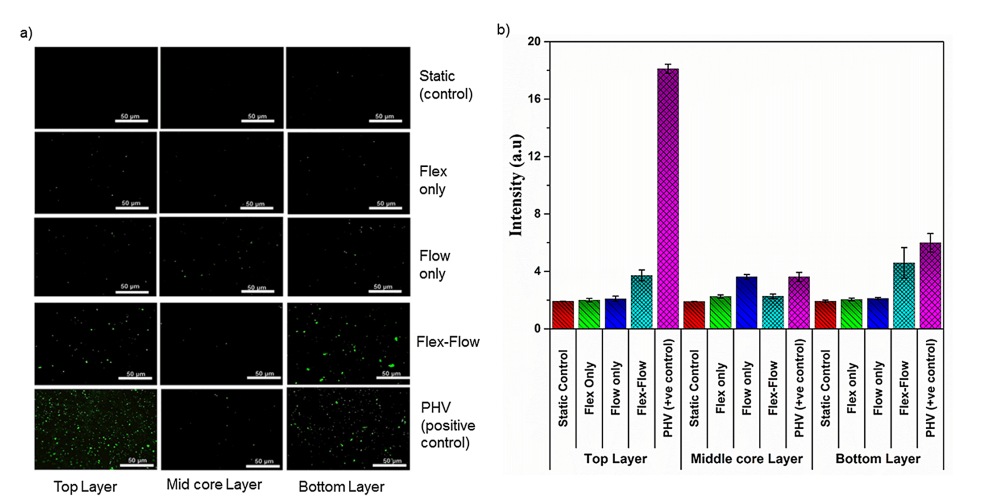
圖 5. EC 標記 CD31 在瓣膜組織的兩個表面層(每側厚度約 90μm)、中間核心部分(厚度約 400 μm)上的免疫熒光染色;第一行:靜態對照;第二行:Flex;第三行:Flow;第 4 行:Flex-Flow 條件;第 5 行:豬心臟瓣膜作為陽性對照。在實驗組中,僅在 flex flow 組中,CD31 表達細胞在工程組織的淺層(頂層和底層)內可見。b) 四個實驗組中 CD31 信號強度的陽性染色(綠色;來自部分 a 中的圖像)的量化;與對照組相比,暴露于 flex-flow 的樣本表達的陽性 CD31 水平明顯更高(p < 0.05)。PHV 被視為陽性對照。
3.4 基因表達
Flex-Flow 條件表明支持瓣膜表型的早期證據:對選定標記物的初步基因表達進行了評估,以確定組合的 FlexFlow 條件是否增強了體外生長結構中的心臟瓣膜表型。
在從 flex-flow 組中提取的 RNA 中觀察到 FZD2 和 YARS 標記物(心血管 SMC(當 MLC1f 未表達時)和 EC 相關基因)的最高表達水平,其中樣品暴露于生物反應器中的組合循環彎曲(1 Hz)和流動(850 ml/min)條件(圖 6);請注意,MLC1f 僅在無流量對照組中顯著表達(P < 0.05)。與研究的其他組(無流量控制、僅流量、僅周期性彎曲)相比,在彎曲流動情況下,對瓣膜發育中的瓣膜形成至關重要的轉錄因子 KLF2A 也顯著表達(p<0.05)(S2 文件)。
圖 6. BMSC 衍生的工程瓣膜組織的基因表達。研究的四個組為:靜態對照、Flow(850 ml/min)、Flex(1 Hz)和 Flex-Flow(同時應用 850 ml/min 流速和 1Hz 頻率對標本進行循環彎曲)。細胞培養基在生物反應器調節室中循環的流速為 850ml/min,允許在內壁和外壁上分別產生 2.91 達因/cm2 和 4.73 達因/cm2 的生理相關 4 流體誘導平均剪切應力。PHV 被視為陽性對照,作為參考。
3.5 計算結果
Flex-Flow 培養標本的剪切應力大小和振蕩屬于心臟瓣膜的生理范圍和剪切應力分布:量化組織工程標本上流體誘導的剪切應力空間和時間分布需要進行 CFD 模擬,然后對結果數據進行分析。
在所有模擬情況下,在內表面和外表面上繪制了流體誘導的時間平均剪切應力大小 (TSSM)(圖 7)。靜態控制、單獨穩定流、單獨循環彎曲和彎曲流動情況下的內壁 TSSM 值分別為 0 達因/厘米2、1.98 ± 0.37 達因/厘米2、0.1 ± 0.005 達因/厘米2 和 2.91 ± 0.11 達因/厘米2。控制、單獨穩定流、單獨循環彎曲和彎曲流動情況下的外壁 TSSM 值分別為 0 達因/厘米2、2.43 ± 0.06 達因/厘米2、0.1 ± 0.003 達因/厘米2 和 4.73 ± 0.09 達因/厘米2。
圖 7. 以下四種情況下,在試樣內壁和外壁上,一個周期內流體誘導的時間平均剪切應力:i) 靜態控制、ii) 流動、iii) 彎曲和 iv) 柔性流動狀態。在比較兩種動態情況 (iii) 與 (iv) 時,柔性流動狀態顯示出更高的剪切應力值。
而外壁(n = 3)的相應值分別為 0.313± 0.011 和 0.094± 0.10。確定面積平均 OSI指標以量化實驗中耦合剪切應力大小和時間振蕩的程度。在樣本區域(樣本大小 n = 3/組),內壁的面積平均 OSI為 0.1±0.05 達因/厘米2(單獨循環彎曲)和 0.41±0.11 達因/厘米2(彎曲流動),而外壁的面積平均 OSI為 0.16 ±0.03 達因/厘米2(單獨循環彎曲)和 0.27±0.16 達因/厘米2(彎曲流動)(圖 8)。請注意,如果流動在時間上是單向的,或者流體引起的剪切應力可以忽略不計,則 OSI- 2 = 0,這分別是單獨穩定流動和無流動的情況。
圖 8. 動態柔性和柔性流動情況下,樣本內外表面的 OSI 縮放剪切應力大小。
四、討論
功能性心臟瓣膜置換,特別是兒科危重瓣膜疾病,迫切需要合適的治療方法;主要限制是,目前可用的置換裝置無法提供軀體生長。因此,自體和活體生物心臟瓣膜替代品的概念非常有吸引力。多項研究表明,開發堅固的工程瓣膜組織的過程需要一個動態培養過程,其中機械應力(例如流動、彎曲和拉伸)會施加到生長的結構上。這些應力狀態尤其重要,因為瓣膜小葉在心動周期中經歷高度變化且依賴于小葉側的流體誘導剪切應力分布,以及局部周期性組織拉伸和彎曲。例如,在主動脈瓣中,與動脈側相比,小葉的心室側經歷的流體誘導剪切應力明顯更高。一些體外研究已經能夠重現這種血液動力學環境的重要特征,使用生物反應器從頭生長瓣膜組織。然而,體外機械調節對結構內細胞分布和表型的具體影響仍不清楚。這些屬性可以支持工程化組織與天然組織的整合,從而可以在指導體內后續瓣膜重塑事件中發揮關鍵作用。此外,維持生理范圍的調節參數(例如流體誘導的剪切應力)的重要性也未得到充分解決。另一方面,迄今為止,干細胞(如 BMSCs)的效用已在幾種心臟瓣膜組織工程方法中得到探索,這些方法已證明在機械刺激環境下組織生長強勁。因此,為了確定瓣膜相關應力環境對干細胞表型的影響,以及機械調節的生理尺度的影響,我們將 BMSC 植入支架置于流體剪切和彎曲應力狀態的單獨和組合模式中,這兩種狀態都與瓣膜組織高度相關。
我們發現 flexflow 組的 BMSC 衍生膠原蛋白含量相對于其他組顯著增加 (p<0.05),而靜態控制、Flex 和 Flow 組之間沒有發現顯著差異 (圖 2)。我們的觀察結果與先前的發現一致,其中亞生理剪切應力下的 flex-flow 研究 (平均壁面剪切應力 = 1.15 達因/厘米2) 與生理水平的周期性彎曲 (1Hz,相當于心率為 60 次/分鐘) 相結合。然而,我們試圖在我們工程組織標本上重現天然主動脈瓣液誘導剪切應力分布的區域變化和側面特異性。事實上,先前的工作已經證明,我們標本的內壁和外壁類似于瓣葉的主動脈側和心室側。在這里,在彎曲流動狀態下,外壁上的 TSSM 為 4.73 達因/厘米2,而沿內壁,剪切應力為 2.91 達因/厘米2。天然人類主動脈瓣的心室側暴露于約 3.87 達因/厘米2 的 TSSM,因此,本研究中確定的彎曲流動標本的剪切應力具有生理相關性。將我們的研究與在亞生理剪切應力水平下進行的研究進行比較,我們發現膠原蛋白的產生額外增加了 70%,即當標本的剪切應力大小和分布在區域上與天然主動脈瓣葉相似時。我們注意到,早期的一項研究通過體外模擬動脈壓力條件證明了生理調節尺度的重要性,最終導致膠原蛋白含量與亞生理流動環境相比增加了約 35%。我們推測,在我們的案例中,差異更為明顯,這是由于細胞調節機制直接導致的,已知這些機制是由流體誘導的剪切應力啟動的,我們認為這些機制也適用于分化 BMSC。
先前的研究已經檢查了 TEHV 中細胞的全厚度分布和表型。與采用亞生理范圍剪切應力調節的柔性流動研究一致,我們觀察到內皮細胞表達主要在表面表達,在間質層中可忽略不計(圖 4)。我們認為,在流動狀態下,BMSC 向內皮細胞的分化和支架內 BMSC 遷移模式得到增強,并且與流量大小沒有很強的依賴性。事實上,我們之前通過磁共振成像可視化并監測了細胞支架遷移模式的增加,發現與無流量控制相比,在穩定流動條件下,這種增加得到增強。另一方面,機械環境與新生瓣膜組織內 α-SMA 表達細胞之間的聯系并不那么簡單。研究人員證明,在受到產生亞生理量級的流體誘導剪切應力的彎曲流動狀態后,表達 α-SMA 的 BMSC 會優先分布在工程心臟瓣膜組織的表面,并表現出強勁表達。同時,Hoestrup 及其同事描述了在生物反應器中受到生理壓力范圍(30 至 75 mmHg)的三葉工程瓣膜結構中表達 α-SMA 的 BMSC 的一致分布;此外,細胞密度在 TEHV 表面最大,而在組織深處則稀疏。我們觀察到,應用心臟瓣膜生理相關的流體誘導剪切應力(TSSM,內壁:2.91dyne/cm2 和外壁:4.73dyne/cm2)有助于促進細胞形成,尤其是表達 αSMA 的 BMSC(圖 4)。雖然這種情況在單獨流動和彎曲流動條件下都會發生,但只有在后者中,α-SMA 表達細胞在工程組織的間質區域內占主導地位,而在表層上則稀疏。另一方面,如前所述,僅在彎曲流動組中,具有內皮表型的 CD31 表達細胞優先分布在工程組織標本的表面上(圖 5)。盡管 α-SMA 和 CD31 免疫熒光染色的程度不如天然瓣膜明顯,正如基因表達結果中瓣膜相關標記物所證明的那樣,很明顯,彎曲流動調節的生理尺度以模仿天然瓣膜細胞分布的方式促進異質細胞分布。我們認為,在體外環境中實現異質性、瓣膜相關的 BMSC 分布和分化的能力在植入后持續組織重塑中起著關鍵作用,包括加速復制原生三層瓣膜結構。這種結構僅在 TEHV 植入后數周(16-32 周)的綿羊研究中觀察到,并且僅限于更換肺動脈瓣,而不是更換要求更高的主動脈瓣或二尖瓣位置。因此,在心臟瓣膜疾病的特定情況下,例如在治療嬰兒嚴重的先天性主動脈瓣狹窄時,加速組織重塑可能特別重要,以跟上軀體生長的步伐。
作為瓣膜形成的進一步證據,我們發現與所有其他組相比,柔性流調節顯著增強了(p < 0.05)KLF2A 基因(圖 6)。在天然瓣膜發育過程中,KLF2A 的缺失會導致嚴重的瓣膜畸形,并直接受到振蕩流體誘導的剪切應力的調節。有趣的是,我們能夠證明,在柔性流動條件下,干細胞(而不是內源性瓣膜細胞,特別是 BMSC)也可能上調 KLF2A 的表達。我們之前的研究表明,OSS 在柔性流動條件下得到增強,因此很可能引發細胞 KLF2A 的上調。
當對 flexflow 組中觀察到的 KLF2A 標記物表達與相應的天然瓣膜基因表達進行集體比較時,我們毫不意外地發現,天然瓣膜表達更高,約為 49%。與從天然瓣膜組織中獲得的細胞的基因表達相比,在時間有限的體外培養環境中,BMSCs 的密度明顯較低,很可能表現出截斷的基因表達水平。然而,需要考慮的更廣泛的論點是,體外瓣膜表型的促進,即使很小,也可能對增強植入后工程化與天然瓣膜組織之間的整合至關重要。從表型的角度來看,周圍體內細胞環境對工程化結構的識別可能對旁分泌細胞信號傳導事件至關重要,這將有助于確保 TEHV 的引導重塑,并最大限度地降低組織過度生長和不受控制的血管翳的風險。先前的研究已經證明了在 TEHV 中重現原生三層組織結構對于支持強大功能的重要性。同樣,我們推測 BMSC 衍生的工程化構造的表型狀態在植入前同樣重要,我們在此表明其針對的是柔性流動機械調節下的瓣膜譜系。因此,我們解釋為在體外培養工程組織期間的柔性流動機械調節以異質方式支持 BMSC 分化,即通過促進瓣膜內皮細胞和間質細胞表型。這一發現證實了我們之前的計算預測,即在柔性流動狀態下同時存在 OSS 和臨界剪切應力會協同增加工程組織中的膠原蛋白含量,這似乎也增強了瓣膜表型,正如這里所證明的那樣。
總之,生理相關的彎曲流動狀態有助于促進體外生長的 BMSC 衍生的工程心臟瓣膜組織中的細胞分布和表型。我們首次能夠證明 BMSC 在工程組織內的分化和遷移導致表面襯里的內皮細胞標志物表達細胞和間質填充的肌成纖維細胞標志物表達細胞,從而類似于天然瓣膜細胞分布。此外,OSS 與臨界水平的剪切應力的共存使得關鍵瓣膜基因(特別是 KLF2A)的非常早期階段能夠強勁表達。請注意,單獨的彎曲狀態會誘導 OSS,但具有可忽略不計的剪切應力大小,并且被發現不會增加工程組織內的細胞和表型;這與我們之前的發現一致,無論是時間方向性還是剪切應力的大小,在引發 BMSC 反應方面都很重要,我們在此展示了這可以在彎曲流動狀態下通過實驗實現。我們得出結論,通過生理相關的彎曲流動條件可以實現 BMSC 衍生的工程化心臟瓣膜細胞組成和表型的體外優化。刺激異質瓣膜細胞和表型的能力可能在指導瓣膜細胞分布的早期階段以及隨后的體內瓣膜組織重塑事件方面很重要,以確保兒科患者 TEHV 的長期成功。
公司地址:廣東省東莞市樟木頭鎮塑金國際1號樓810
責任編輯:
Copyright ? 2022 數耕包裝 本站部分資源來源于互聯網 我們愿與各界朋友,共建環保未來! 蜀ICP備2023018497號-1 XML地圖
